بر اساس قانون مادری در بیولوژی مولکولی یا Central Dogma، که اولین بار در سال 1958 توسط آقای فرانسیس کریک مطرح گردید، ماده اصلی ژنتیکی DNA میباشد که از روی آن mRNA سنتز شده و mRNA در نهایت به پروتئین ترجمه میشود (هرچند از لحاظ تکاملی و فیلوژنی RNA زودتر از DNA بوجود آمده است). سلول قادر است تولید یک پروتئین را در سطح رونویسی یا ترجمه کنترل کند. مکانیسم مذکور، مکانیسم تنظیم بیان ژن نامیده میشود. در این فرآیند تنظیمی:
- برخی از پروتئینها مثل فاکتورهای رونویسی با اتصال به پروموتور، افزایشگر و یا خاموشگرهای یک ژن، رونویسی از آنها را متاثر می کنند (مثل فاکتور GATA-1 که با اتصال به پروموتور ژن EOS47 باعث افزایش بیان آن و تمایز ائوزینوفیلی میشود یا دایمر Maf-Bach که با اتصال به خاموشگر ژن گلوبین باعث مهار رونویسی آن می گردد).
- برخی از پروتئینها مثل HATCEP/P300، HDAC و متیل ترانسفراز با تغییرات اپیژنتیکی بر روی DNA، هیستونها و با تغییر در برهمکنش DNA با پروتئینهای هیستونی، رونویسی از یک ژن را تحت تأثیر قرار میدهند. تغییرات اپیژنتیکی برخلاف تغییرات ژنتیکی، توالی نوکلئوتیدهای DNA را تغییر نداده ولی با متیلاسیون بازهای سیتوزین یا با استیلاسیون، داستیلاسیون، متیلاسیون و فسفریلاسیون هیستونها، ساختار کروماتین را به نوعی تغییر میدهند که شرایط ساختاری و فیزیکی برای رونویسی از یک ژن کاهش یا افزایش یابد.
- برخی از پروتئینها با اتصال به فاکتورهای رونویسی، عملکرد آنها را بلوکه کرده، باعث کاهش برهمکنش آنها با DNA شده و بدین ترتیب رونویسی از ژنهای هدف آنها را کاهش میدهند (همانند پروتئین IKB که باعث مهار NF-kB میشود). برعکس برخی از پروتئینها نیز با اتصال به فاکتورهای مهارکننده رونویسی و بلوکه کردن آنها، سبب حذف اثر مهاری و افزایش بیان ژنهای هدف میگردند (همانند IKK که با مهار IKB باعث افزایش فعالیت NF-kB و افزایش رونویسی از ژنهای هدف میشود).
- برخی از پروتئینها به واسطه سیگنالینگ داخل سلولی و ایجاد توانایی ورود یک فاکتور رونویسی از سیتوپلاسم به داخل هسته، بیان یک ژن را افزایش می دهند (همانند دایمر STAT که طی مسیر سیگنالینگ JAK-STAT فعال شده و با ورود به هسته باعث رونویسی از ژنهای هدف میشود).
- برخی از پروتئینها با اتصال به زیرواحدهای مختلف ریبوزوم باعث جلوگیری از ترجمه یک mRNA و کاهش بیان یک ژن میشوند.
- برخی از پروتئینها با اتصال به پروتئینهای دخیل در شروع یا طویل سازی ترجمه باعث کاهش سطح ترجمه و درنتیجه کاهش بیان یک ژن میشوند (همانند فاکتور HRI که با اتصال به eIF2 سبب فسفریلاسیون و غیرفعالسازی آن و درنتیجه کاهش سطح ترجمه میشود).
- برخی از پروتئینها با اتصال به یک پروتئین و یوبیکوئیتینه کردن آن، سبب تخریب زودرس و کاهش بیان آن میشوند (همانند پروتئین β-TrCP که با یوبیکوئیتیناسیون β-کاتنین باعث تخریب آن در پروتئوزوم شده و از عملکرد آن جلوگیری میکند یا پروتئین VHL که با تخریب HIF-α مانع از سیگنالینگ آن میشود).
- برخی از پروتئینها با اتصال به نواحی 5′ یک mRNA مانع از اتصال آن به ریبوزوم و کاهش سطح ترجمه میشوند (همانند پروتئین IRP که با اتصال به mRNA فریتین باعث کاهش سطح ترجمه آن میگردد).
- برخی از پروتئینها با اتصال به نواحی 3′ یک mRNA باعث افزایش پایداری mRNA و افزایش سطح ترجمه میشوند (همانند پروتئین IRP که با اتصال به mRNA گیرنده ترانسفرین یا CD71 سبب افزایش سطح ترجمه آن میشود).
- برخی از پروتئینها با اتصال به RNAهای کوچک غیرکدکننده به یک کمپلکس ریبونوکلئوپروتئینی تبدیل میشوند که از طریق RNA داخل آن به mRNAهای سیتوپلاسمی متصل شده و باعث برش آنزیمی و تخریب mRNA و درنتیجه کاهش ترجمه و بیان آنها میشوند. از این دسته از RNAها میتوان به RNAiها (miRNAها، SiRNAها و piRNAها) اشاره نمود که در ادامه در مورد ساختار و عملکرد اختصاصی آنها بحث خواهد شد.
- برخی از mRNA ها مثل ریبوسوئیچ، در صورت اتصال به مواد متابولیت باعث کاهش ترجمه mRNA موجود در ریبوسوئیچ میشوند.
تا سالهای طولانی تصور بر این بود که برای هر پروتئینی یک ژن وجود دارد (قانون یک ژن یک پروتئین)، ولی بعدها با کشف ایزوفرمهای مختلف از یک پروتئین و با شناسایی پدیده پردازش تمایزی مشخص شد که از یک ژن و بسته به آرایش اگزونهای مختلف آن، ایزوفرمهای متفاوتی از یک پروتئین میتواند شکل گیرد که حتی محل بیان آن نیز ممکن است از یک سلول به سلول دیگر یا از یک بافت به بافت دیگر فرق داشته باشد. از طرفی دیگر، تعداد پروتئینهای موجود در انسان به مراتب بیشتر از تعداد ژنهای موجود در کل 46 کروموزوم آن میباشد که این یافته نیز تشکیل چندین پروتئین از یک ژن را تایید می نماید. طی فرآیند بلوغ hn-RNA به mRNAی بالغ، اینترونهای hn-RNA حذف شده و اگزونها به یکدیگر متصل میشوند ولی طی پردازش تمایزی، ترکیبات متنوعی از اگزونها ایجاد میشود که بسته به نوع آن، پروتئینی با عملکرد، بیان، ساختار، طول پپتیدی و دومینهای متفاوت ایجاد میگردد.
از کل ژنوم انسانی تنها، حدود 5% آن کد شده و به پروتئین تبدیل میشود و 95% بقیه، بیشتر به صورت هتروکروماتین و ساختارهای اپیژنتیکی قرار دارد که در حفظ ساختار DNA، توپولوژی آن، تقسیم میتوز و میوز و مهمتر از همه، در تنظیم بیان ژنها ایفای نقش میکند. چنین عنوان میشود که در هر10% از یک کروموزوم، حدود 40 ژن وجود دارد (4 ژن برای 1% از کروموزوم)، حال اگر کروموزومی به طور متوسط 106×48-22 جفت باز داشته باشد، به ازاء هر 104×4/3 باز، بهطور متوسط یک ژن وجود خواهد داشت. لازم به ذکر است، کل DNA انسان حدود 3 میلیارد جفت باز دارد که در 46 کروموزوم توزیع شده است و در این تعداد باز، حدود 35-30 هزار ژن وجود دارند. بیش از 50% DNA هم از توالیهای تکراری تشکیل شده است. در خود mRNA ای که از یک ژن رونویسی میشود نیز، نواحی ترجمه شونده و غیرترجمه شونده (UTR) وجود دارد که نواحی UTR (مثل اینترونها)، در تولید پروتئین دخالت نداشته و حین بلوغ mRNA، از آن جدا میشوند. امروزه مشخص شده که اینترونها در تولید مقدار قابل توجهی از miRNAها حائز اهمیت هستند.
ژنهای کد شونده دو دسته هستند، برخی از آنها (مثل اکتین، آنزیمهای گلیکولیز و ژن سازنده ATP)، در تمامی سلولها و تمامی مقاطع تکاملی بیان شده و بهعنوان مایحتاج روزانه و خانگی یک سلول به شمار میروند و ژنهای خانه دار یا house kipping نامیده می شوند. این ژنها تنظیم بیان خاص و پیچیدهای نداشته و به راحتی در شرایط مختلف سلول بیان میشوند. در مقابل ژنهایی وجود دارند که مختص بافت و سلولهای خاصی بوده و یا در مقطع تکاملی خاصی بیان میشوند (مثل ژن تستسترون در بیضه یا ژن سازنده انسولین در پانکراس) که دارای تنظیم بیان ژنی مشخص و زمانبندی شدهای بوده و ژنهای خاص نامیده میشوند.
RNA های کدکننده، mRNA ها هستند، و RNAهای غیرکدکننده(nc-RNAs) شامل موارد زیر میباشند:
- tRNA(transfer RNA)
- rRNA(ribosomal RNA)
- snRNA(Small nuclear RNA- splicing)
- snoRNA(Smal nucleolar RNA)
- Natural Antisense یا RNA interference (RNAi)
- miRNA یا microRNA
- Ribozyme
- Riboswitch
- siRNA
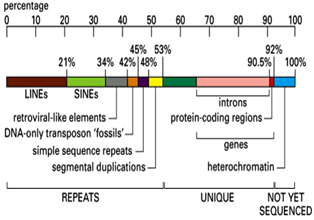
نسبت قسمتهای مختلف DNAی انسانی که حدود 95% آن را هتروکروماتین و 5% آن را یوکروماتین تشکیل میدهد. از کل کروموزومها،30% آن را ژنها تشکیل میدهد که از این 30%، 5/1% آن اگزون و 5/28% مابقی آن اینترونهای غیرکدکننده هستند. از نگاهی دیگر، حدود 53% DNA انسان از توالیهای تکراری و 5% آن از نواحی مضاعف شده تشکیل می گردد و حدود 8% از ژنوم انسان نیز منشاء رتروویروسی دارد.

snRNAها گروهی ازRNA ها هستند که تنها در سلولهای یوکاریوتی دیده میشوند و توسط RNA پلیمراز IIیا III کد میگردند. این مولکولها در فرایندهای مهمی همانند پردازش، تنظیم فاکتورهای رونویسی، تنظیم RNA پلیمراز II و نگهداری تلومر نقش دارند. snRNA ها را به همراه پروتئینهای همکارشان، اسنورپ نیز مینامند. SnoRNAها، دستهای از snRNAها هستند که در بیوسنتز RNA و راهنمایی تغییرات شیمیایی مانند متیلاسیون درtRNA، rRNA و snRNA دخالت دارند. این دسته در هستک و در بخشی از هسته به نام Cajal body که محل سنتز RNA میباشد، قرار دارند. انواع non-coding RNA بسته به عملکردی که دارند، به انواع مختلفی تقسیم میشوند، بهعنوان مثال دستهای از آنها در مهار رونویسی نقش دارند (miRNA, Antisense)، دستهای در تخریب (siRNA, Ribozyme, Antisense) RNA ، دستهای در بازسازی کروماتین (dsRNA)، دستهای در متیلاسیونDNA (dsRNA)، دستهای در Imprinting ژنی (non-coding antisense)، دستهای در پردازش hnRNA نابالغ (snRNA)، دستهای در Editingو ترمیم (snoRNA)، دستهای در رونویسی معکوس (telomerase, retrotransposone)، دستهای بهعنوان سنسورهای متابولیکی (Riboswitch) و دسته ای در تعدیل عملکرد پروتئینها ((OxyS نقش دارند. به فرمهای سنتتیک Antisenseها مورفولینو نیز گفته میشود.